« El Universo y la Mente »
-

-
 Descarga gratuita
Descarga gratuita
Emilio Silvera Vázquez
« Imagen del Día »

« Colaboraciones »
-
La cuántica fácil
Fandila Soria Martínez -
Física global
Germán Vidal -
Inteligencia extrema
Germán Vidal -
Cosmología no convencional
Ramón Marqués -
Modelo Cosmológico 2017
Germán Vidal -
Monopolos Gravitacionales
Germán Vidal -
Física del Todo
Germán Vidal -
Física del Todo. Capítulo 33
Germán Vidal -
 Inocentes como ellos
Inocentes como ellos
Fandila Soria Martínez -
J E N A R O
Fandila Soria Martínez -
La caverna
Fandila Soria Martínez -
Los inciertos frutos
Fandila Soria Martínez -
Tres cuentos y uno más
Fandila Soria Martínez -
Un artístico triángulo
Fandila Soria Martínez -
Una cuántica razonableTemporalmente en revisión
Fandila Soria Martínez -
Agujeros Negros, origen y dinámica relativista
Germán Vidal -
Fase de ruptura ambiental
Germán Vidal -
Procedimiento FRP
Germán Vidal -
Folleto oficial de FRPV
Germán Vidal -
Alerta de calentamiento global
Germán Vidal -
Teoría de la planificación universal
Dante Pracilio
« Escrito recientemente »
- El fascinante “universo” de las partículas
- Los grandes números distintivos del Universo
- Se acerca el verano, época de vacaciones
- ¡Ojo! China avanza calladamente, pero…
- Desde la materia “inerte” hasta los pensamientos
- Imaginación sin límite pero… ¿Sabremos comprender?
- ¡El Universo! Y nosotros… ¿Seremos su parte que piensa?
- Seguimos sin querer enterarnos
- ¿El viaje a otras estrellas? Un engaño de la mente para evitar la frustración
- Nadie pudo escribir, la Historia de la Vida
Últimos comentarios
- en ¿Es la “Estructura” más compleja del Universo?
- en Puede la I.A., ser un peligro para la Humanidad?
- en Los cuasicristales: un nuevo orden de la materia
- en Los cuasicristales: un nuevo orden de la materia
- en Enigmático Encélado
- en Enigmático Encélado
- en La Física de Partículas al Servicio de la Salud
- en Necesitamos una teoría unificada del Cosmos
- en El placer de Descubrir: Aventurarse por nuevos caminos.
- en El placer de Descubrir: Aventurarse por nuevos caminos.
« Categorías »
- 30 Millones de visitas!
 (1)
(1)
- a (1)
- a otros mundos (18)
- a pesar de todo (4)
- Acelerador en Huelva (1)
- Aceleradores de partículas futuros (4)
- Agujeros de gusano (1)
- Agujeros negros (69)
- AIA-IYA2009 (206)
- Albert Einstein (3)
- Alcanzar otra dimensión (2)
- Algo de Cine (2)
- Algo de lo que pasó desde el Big Bang (8)
- Alquimia (10)
- Alquimia estelar (31)
- Ancestros (1)
- Andrómeda (2)
- Anécdotas de personajes de la Ciencia (6)
- Antimateria (8)
- Aquella cancioncilla (1)
- Aquellos filósofos de la naturaleza (3)
- Aquellos genios (3)
- Artículo de Prensa (9)
- Así etán las cosas (9)
- Asteroides (4)
- Astrofísica (54)
- Astronomía y Astrofísica (571)
- Avances hacia el futuro (6)
- Bacterias nosivas (1)
- Belleza sí (5)
- Big Bang (1)
- Biologia (96)
- Bioquímica (31)
- Breve historia del Universo (1)
- Burlar la Velocidad de la Luz (1)
- Cambios inesperados (1)
- Canción de desamor (2)
- Caos y Complejidad (11)
- Carnaval de Física (4)
- Carnaval de Matematicas (15)
- Catástrofes Naturales (83)
- Causalidad… Ese Principio (3)
- Celebraciones (5)
- Cerebro y Mente (13)
- Ciencia futura (49)
- Ciencia y Pseudociencia (1)
- Ciencia y religión (3)
- Ciencia y Vida (16)
- Ciencias de la Tierra (13)
- Civilizaciones antiguas (10)
- Colaboración (6)
- Colaboraciones (2)
- Comentario a la imagen del día (4)
- Computación cuantica (1)
- conciencia (28)
- Conferencia (2)
- Conjeturas (5)
- Conocer el Universo (3)
- Conocer la Naturaleza (3)
- Conociendo el Sistema Solar (5)
- Constantes universales (20)
- Contaminación radiactiva (1)
- Cosas curiosas (46)
- Cosas que no deben pasar (2)
- Cosas que pasan (5)
- Cosmología (43)
- Cosmología de los Antiguos pueblos (4)
- Cosmología de vacío (1)
- Curiosidades (31)
- De estrella a púlsar (2)
- De lo pequeño a lo grande (4)
- Debates (20)
- Dehumanizados (1)
- del pasado al presente (1)
- Densidad Crítica (8)
- Descubriendo secretos del Universo (10)
- Descubrir y aprender (39)
- desde la materia inerte hasta los pensamientos (5)
- Desde la materia inerte ¡Hasta los pensamientos! (4)
- Deseos que nunca serán cumplidos (1)
- Dignidad (1)
- Divagando (64)
- Diversidad (8)
- Divulgando la ciencia (5)
- Divulgar la Ciencia (1)
- Ecos del Big Bang (5)
- El "universo de las galaxias" (2)
- El "universo" de la Consciencia (5)
- El "universo" de los Fractales (1)
- El agua de Marte (1)
- El agua… ¡esa maravilla! (3)
- El Agua: Un tesoro para la vida (1)
- El Amor (4)
- El Arte (5)
- El Carbono (1)
- El Centro Galáctico (1)
- El cerebro (32)
- El CERN (2)
- El cielño en febrero/2015 (1)
- El comentario del visitante (2)
- El Cuerpo Humano (3)
- El Destino… Esa variable (1)
- EL DETERIORO DEL PLANETA tIOERRA (1)
- El divagar de la Mente (13)
- El Espacio Exterior y nosotros (11)
- El fin del Universo (2)
- El Final del ciclo solar (3)
- el futuro (41)
- El Futuro incierto (75)
- El futuro tecnológico (14)
- El hombre en el Universo (31)
- El Invento del Alma (2)
- El libre pensamiento (4)
- El maldito dinero (1)
- El Medio Ambiente (1)
- El mejor amigo: Un libro (2)
- El Metano Marciano (2)
- El misterio de las Galaxias que se destruyen (1)
- El misterioso número Pi (2)
- El Modelo Estánfar (5)
- el Mundo y nosotros (23)
- El núclo atómico (6)
- El origen (23)
- El Origen de las cosas (21)
- El origen de los elementos (5)
- El pasado (6)
- El pasado nunca volvera (1)
- El placer de descubrir (3)
- El Planeta Tierra (1)
- El Plasma: cuarto estado de la materia (3)
- el presente y el futuro incierto. (3)
- El primer contacto (6)
- El saber del mundo (27)
- El saber ocupa lugar y tiempo (2)
- El saber: ¡Ese viaje interminable! (44)
- El Ser consciente (2)
- El Sistema Saturno (1)
- El Sistema Solar (9)
- El Tiempo inexorable (9)
- El Tiempo pasa…¿O somos nosotros? (14)
- El Tiempo siempre presente (4)
- El Universo (80)
- El Universo asombroso (394)
- El Universo cambiante (37)
- El Universo de Ayar y el Universo de Hoy (10)
- El Universo de la Conciencia (18)
- El Universo dinámico (157)
- El Universo Hiperdimensional (15)
- El Universo misterioso (206)
- El Universo y la Entropía (22)
- El Universo y la Mente (74)
- El Universo y la Química de la Vida (89)
- El Universo y la Vida (324)
- El Universo y los pensamientos (67)
- El Universo y… ¿nosotros? (217)
- El Universo: Todo Energía (21)
- elementos (1)
- En nuestro Universo: La Eternidad no existe (1)
- Encuentros Espaciales (1)
- Encuesta (1)
- Energía = Materia (9)
- Energia de fusión (2)
- Energías de la Tierra (8)
- Enigmas del Corazón (2)
- enigmas por resolver (5)
- Entrevista (4)
- Entrevista científica (10)
- Entropía (5)
- Es bueno recordar lo que pasó (4)
- Esa Ilusión llamada ¡Tiempo! (1)
- ese misterio (5)
- Especulando (5)
- Este mundo injusto (2)
- Estrellas (22)
- Estrellas de neutrones y Púlsares (2)
- Estrellas fugaces (1)
- Estrellas masivas (4)
- Estructuras fundamentales (6)
- Eternidad? (1)
- Eva mitocondrial (1)
- evadirnos (1)
- Evadirnos del mundo por un momento (1)
- Eventos (2)
- Evolución (15)
- Experimentos espaciales (1)
- Exploración de los mundos (2)
- exploración del espacio (4)
- explorando lo "nano" (1)
- Extinciones (4)
- Extinciones de origen desconocido (1)
- Extraño sueño (1)
- Fallida Misión a Marte (1)
- Felicidad para todos (1)
- Felicitar a un amigo (1)
- Fenómenos naturales (1)
- Ficción (1)
- Filosofía (7)
- Fin de la Misión Cassini (1)
- Física (1.650)
- Física Cuántica (517)
- Física Relativista (68)
- La Mujer en la Ciencia (2)
- Física Cuántica
- Física de vacío (9)
- Física en las estrellas (2)
- Física Solar (1)
- Física y cosmología (6)
- Física-química (16)
- Física… ¡Y mucho más! (20)
- Formación de elementos (2)
- Fuerzas de la Naturaleza (3)
- Fusión de galaxias (1)
- Futuro (17)
- Gaia (12)
- Gases nobles (1)
- General (4.015)
- Genética (1)
- Gracias al visitante (1)
- Grandes extinciones (1)
- Hacia el futuro (21)
- Hacienda somos todos (2)
- Hay que sentir (1)
- Hiperespacio (2)
- Historia para mirar (6)
- Humanidad (16)
- I. A. (15)
- Imaginación (19)
- Imaginando (2)
- Implosión de una estrella (1)
- Injusticia sin fin (3)
- Inocentada (1)
- Interacciones fundamentales (2)
- Internet (1)
- Investigación y Ciencia (3)
- La Astronomía y la Humanidad (4)
- La Belleza (1)
- La Belleza y la Ciencia (5)
- La Ciencia (6)
- La Ciencia debe avanzar (2)
- La Ciencia en el pasado (2)
- La Ciencia Fiscción (1)
- La Complejidad (9)
- La complejidad de la Vida (12)
- La Conquista del Espacio (4)
- La contaminaciñon del planeta (1)
- La Entropía lo destruye todo (6)
- la Entropía siempre presente (2)
- La estructura del Espacio (1)
- La física en la vida cotidiana (2)
- La Física y el Universo (1)
- La Física y la Salud (2)
- La formación de las galaxias (2)
- La fotosíntesis (1)
- La fragilidad HUmana (1)
- La Geotectónica (1)
- La ignorancia nos acompaña siempre (30)
- La Imagen del día (1)
- La Implosión de las estrellas (1)
- La importancia del tiempo (1)
- La inmortalidad no existe (1)
- La justa medida (5)
- La libertad de pensar (2)
- La luna Europa (1)
- La Luz esconde muchos secretos (13)
- La magia de la Tierra (6)
- La mágica Naturaleza (4)
- La maldad Humana (1)
- La materia tiene memoria (5)
- la mente (3)
- La Mente – Filosofía (135)
- La muerte de las estrellas (1)
- La muerte del Sol (1)
- La mujer y la Ciencia (1)
- La música son sentimientos (6)
- La músuca. (2)
- La NASA (1)
- La NASA experimenta (1)
- La naturaleza de la Luz (1)
- La Naturaleza ¡Es sabia! (7)
- La Naturaleza…El Universo (39)
- La Oración (2)
- La realidad cambiante (7)
- La realidad humana ¿es realidad? (16)
- la realidad presente (2)
- La Teoría de Cuerdas (6)
- La Tierra se recicla (2)
- La Tierra y su energía (20)
- La Tierra: Pasado y futuro (1)
- La Unión hace la fuerza (1)
- La vecindad galáctica (3)
- La Vía Láctea (1)
- La vida (20)
- La Vida de las Partículas (1)
- La Vida en la Tierra (4)
- La Vida en otros mundos (4)
- La vida sigue (3)
- Las bacterias y nosotros (1)
- Las constantes de la Naturaleza (9)
- las constantes y la Vida (2)
- Las constantes y las inesperadas (2)
- Las distancias en el Espacio (6)
- Las ecuaciones (3)
- las estrellas y la Vida (20)
- Las galaxias generan entropía negativa (3)
- Las huellas del pasado (8)
- Las nuevas tecnologías (1)
- Las prodigiosas ideas (2)
- Las religiones (1)
- Libre Albedrío (1)
- Lo que creemos que sabemos (7)
- lo que es (4)
- Lo que no sabemos (24)
- lo que será (1)
- Los 100 descubrimientos más grandes (1)
- Los cráteres de la Tierra (1)
- Los Elementos (3)
- Los estados de la materia (3)
- Los Ingleses en Huelva (1)
- Los misterios del Universo (8)
- Los Pensamientos (28)
- Los pilares del Sol en la Tierra (1)
- Los primeros pasos (2)
- Los recuerdos (2)
- Los secretos del Universo (5)
- Los sentimientos (4)
- Los terremotos (1)
- Lunas misteriosas (5)
- malos tiempos (1)
- Maravillosa Teoría (3)
- Marte (81)
- Matemáticas (6)
- Materia extraña (11)
- Materiales increibles (5)
- Materias diversas (4)
- Mecánica cuántica (8)
- Memorias del pasado (1)
- Meteoritos (3)
- Meteoritos asesinos (3)
- Mi hija María (2)
- Mi Huelva (1)
- Mi Tierra (4)
- Mirar el futuro con los pies en el suelo (1)
- Misterios de Júpiter (1)
- Misterios de la Mente (3)
- Misterios del Universo (8)
- Misterios sin resolver (13)
- Moléculas precursoras de la vida (5)
- Mujer y hombre: 2 caras de la misma moneda (1)
- Mujeres científicas (1)
- Multiverso (16)
- Mundo Futuro (6)
- Nada muere y todo cambia (2)
- nada permanece (6)
- Nada puede surgir de la Nada (1)
- Nanotecnología (8)
- Naturaleza (19)
- Naturaleza misteriosa (36)
- Naturaleza-Imaginación (3)
- Nebulosas (23)
- Nebulosas y estrellas (11)
- Newton (2)
- Niburu El Planeta X (1)
- no lo comprendo! (1)
- No solo de pan vive el hombre (6)
- No todo es Ciencia (1)
- nosotros (1)
- Nostalgia (1)
- Noticia comentada (20)
- Noticias (154)
- Noticias NASA (2)
- Nuestra Salud (1)
- Nuestro entorno (5)
- Nuestro entorno…Nuestro futuro (6)
- nuestro entrañable hermano (1)
- Nuestro increíble planeta (1)
- Nuestro origen (1)
- Nuestro recorrido histórico (1)
- Nuevas posibilidades (3)
- Nuevas revoluciones científicas (2)
- Nuevos materiales (4)
- Nuevos mundos (8)
- números adimensionales (1)
- Nunca dejaremos de hacer preguntas (3)
- Omega Negro (1)
- Ondas gravitacionales (19)
- Origen de las matemáticas (1)
- Orion (2)
- Otras clases de vida (3)
- otras formas de vida (4)
- Otras teorias (1)
- Otros mundos (25)
- Panspermia (1)
- Paradojas de la relatividad (2)
- Pasado que sigue siendo presente (1)
- Pensamientos (8)
- Pequeño reportaje (2)
- pero Eternidad… (1)
- Personajes de la Historia (11)
- Personajes ilustres (5)
- Planetas imposibles para la vida (2)
- Preguntas que no sabemos contestar (1)
- presente y futuro de la Ciencia (2)
- Pueblos de Huelva (1)
- Púlsares y galaxias (6)
- Querencias (2)
- Queriendo saber (12)
- Química (33)
- Química estelar y Vida (2)
- Radiación Cósmica (2)
- realidad mañana (1)
- Recordando el pasado (9)
- Recordar la vijeo y querer lo nuevo (1)
- Refinando teorías (1)
- Relación del Sol con la Tierra (3)
- Relatiovidad Especial (3)
- Rememorando el pasado (8)
- Reportajes de prensa (10)
- Resumen del año (2)
- Rotura de la simetrísa CP (1)
- RSEF (1)
- Rumores del Saber (251)
- Rumores del saber del mundo (34)
- sabremos (3)
- Saturno (1)
- Se puede traspasar la Memoria? (1)
- Secretos del Universo (1)
- Seguimos elucubrando (1)
- Siempre hacienda pregutas (1)
- siempre misteriosa (2)
- Simetrías (22)
- Singularidad (2)
- Son tantas las cosas que no sabemos (2)
- Sueños de hoy (1)
- Sueños de la Humanidad (4)
- Superconductores y el campo de Higgs (2)
- Supergravedad (2)
- Supernova (1)
- Sustancias (1)
- También los planetas evolucionan (1)
- Tecnología futura (2)
- Tenemos que saber (6)
- Teoría de cuerdas y dimensiones extra (4)
- Teoría de Supercuerdas (8)
- Teorías ¿Imposibles? (2)
- Teorísa del Todo (1)
- Titán (4)
- todo es número (1)
- Transiciones de fase (3)
- Transiciónes de fase en las estrellas (1)
- Un desahogo (2)
- Un Genio Matemático (1)
- Un Mundo justo (1)
- Un mundo mejor (2)
- Un recorrido desde el comienzo del tiempo (2)
- Un Universo con dimensiones extra (1)
- Una pequeña entrevista (1)
- Unificar la Naturaleza (1)
- universo (1)
- Universo de fantasías (1)
- Universo estacionario y elementos (1)
- Universo primitivo (1)
- Universos paralelos (18)
- Vía Láctea (1)
- Viajar al Espacio (13)
- Viajar al pasado (15)
- vida (1)
- Vida en otros mundos (24)
- WIMPs (1)
- ¡Asombroso! (2)
- ¡Cosas del Universo! (3)
- ¡El futuro es imparable! (1)
- ¡El maldito dinero! (1)
- ¡El Tiempo! ¿Qué será? (1)
- ¡El Tiempo! ¿Qués es el Tiempo? (8)
- ¡Energías! (1)
- ¡Humanidad! (8)
- ¡Imaginación! (7)
- ¡Indignados! (1)
- ¡La amistad! (1)
- ¡La Ciencia! esa maravilla (1)
- ¡La Curiosidad! (1)
- ¡La Gravedad! Esa fuerza misteriosa (1)
- ¡La Humanidad! (2)
- ¡La Materia Oscura! (2)
- ¡La Mente! Ese prodigio (5)
- ¡La Mujer! ¿Cuando reconoceremos su valía? (1)
- ¡La música nos hace mejores! (1)
- ¡La vida! El misterio persiste (6)
- ¡Las estrellas! (3)
- ¡Las matemáticas! (1)
- ¡Los pensamientos! (3)
- ¡Los pensamientos! ¿quién los sujeta? (1)
- ¡Maldita desigualdad! (1)
- ¡Males del mundo! (2)
- ¡Maravillas del Universo! (1)
- ¡Naturaleza! (1)
- ¡Necesitamos saber! (6)
- ¡No estamos sólos! (3)
- ¡NO! (1)
- ¡Noticias! (9)
- ¡Partículas! (1)
- ¡Pueblos y lugares! (1)
- ¡Qué cosas! (2)
- ¡Qué mundo este nuestro! (1)
- ¡Tenemos que saber! (12)
- ¡Tiempo! (3)
- ¡Viajar en el Tiempo! ¿Podremos? (5)
- ¡vIAJES EN EL tIEMPO! (1)
- ¡¡La Ciencia!! (2)
- ¿Alma inmortal? (1)
- ¿Biofísica? (1)
- ¿Cómo se formaron las galaxias? (1)
- ¿Cuánta materia hay en el Universo? (1)
- ¿Cuánta materia vemos? (1)
- ¿De dónde venimos? (1)
- ¿De donde venimos? ¿Quiénes somos? (1)
- ¿Dónde estamos dentro de la Galaxia? (1)
- ¿El primer contacto? ¡Tndrá que esperar! (1)
- ¿El Universo? ¡Tenemos que conocerlo mejor! (2)
- ¿Estrellas de Quarks? (1)
- ¿Extraterrestres? (1)
- ¿Hacia dónde nos lleva la ignorancia? (1)
- ¿Igualdad? ¿Dónde? (1)
- ¿La materia Oscura! (2)
- ¿Libre albedrío? (1)
- ¿Materia oscura? (1)
- ¿Materia Oscura? ¿Dónde? (1)
- ¿Mensajes del futuro? (1)
- ¿Multiverso? (2)
- ¿Mutación? (1)
- ¿Observadores del Universo! (2)
- ¿Ondas gravitacionales? (1)
- ¿otra clase de materia? (1)
- ¿Otros Universos? (2)
- ¿Panspermia? (1)
- ¿Qué es el Tiempo? (1)
- ¿Qué es en realidad la Luz? (1)
- ¿Qué es la belleza? No es lo mismo para todos (1)
- ¿qué sorpresa nos dará? (1)
- ¿Quiénes somos? (1)
- ¿Recordar u olvidar? Qué será mejor (1)
« Archivo »
« Enlaces internos »
- Facebook de ESV
- Amigos de la Física E/hc
- Artículos
- Contacto
- Currículum Vitae
- Galería de imágenes
- Glosario de términos
- Libretas
- Web de Emilio Silvera Vázquez
- Recursos para su web
« Enlaces »
- Asociación Huelva Nueva York
- Carnaval de Física
- Ciencia Kanija
- El Tamiz
- Escritores
- Gravedad Cero
- HIRISE
- NovaCiencia
- Off-Topic Observatorio
- Real Sociedad Española de Física
- ¡Maldito Capital! - Se describe lo que es la realidad del mundo
« Administración »
« Buscar »
« Suscripciones web »










Geolocalizador
« Visitas »
 Totales: 89.951.820
Totales: 89.951.820 Conectados: 145
Conectados: 145
Feb
15
Marte: cada día menos misterioso
 por Emilio Silvera ~
por Emilio Silvera ~
 Clasificado en Marte ~
Clasificado en Marte ~
 Comments (0)
Comments (0)





Nuestro planeta, la Tierra, forma parte del Universo, y, es una prueba indiscutible de que sus componentes biológicos y físicos forman parte de una única red que funciona de un modo autorregulado, y, de esa forma, mantiene las condiciones que son ampliamente adecuadas para la existencia de vida, pero que sufren fluctuaciones a todas las escalas (incluidos los ritmos de alternancia de glaciaciones y periodos interglaciales, así como las extinciones masivas). En un sentido real, la Tierra es el lugar que alberga una red de vida, y la existencia de esta red (Gaia) sería visible para cualquier forma de vida inteligente que hubiera en cualquier otro planeta y que fuera capaz de aplicar la prueba conocida de Lovelock y buscar señales de reducción de la entropía.
Ni la NASA, se tomó nunca la prueba de Lovelock lo suficientemente en serio como para aplicarla a la búsqueda de vida en el Sistema Solar; pero si se lo tomó en serio para buscar vida más allá del Sistema Solar. Claro que después, parece que recapacitaron y enviaron al planeta Marte ingenios robotizados que forman ya una larga lista y, cada una de esas misiones, nos posibilitaron conocer mejor aquel planeta. Se encontró hielo de agua, han diluido porciones de la tierra marciana en agua y debidamente tratada, han hallado la presencia de magnesio, sodio, potasio y cloruros. Sabemos, por las maravillosas fotografías allí tomadas, que el planeta conoció mejores tiempos en los que, mares y océanos y grandes conterrías de agua, eran lo natural y hemos podido ver las huellas de todo aquellos maravillosos procesos. Ahora tenemos más que evidencias de la presencia del agua, la presencia de sales y otros vestigios, así lo confirman y, los compuestos para la vida…andan cerca.

Se están analizando los gases y los compuestos químicos del suelo y del hielo allí encontrados, y, todo ello, debidamente procesado nos dará una respuesta de lo que allí existe. La Curiosity está haciendo su trabajo y hará posible que los datos sobre aquel planeta sean cada vez más abundantes y fiables.

El Invernadero Marciano
En muchas de las fotografías que nuestros ingenios espaciales han tomado de muchos lugares de Marte, podemos ver, sin ningún lugar a dudas, que el agua corrió rumorosa por sus regajos y canales. Los “ríos marcianos” ofrecen una clara evidencia de que el planeta fue en algún momento más caliente y más húmedo. Pero, ¿cómo pudo ser esto? A primera vista hay una buena razón para creer que Marte debería haber estado aún más frío en el pasado que lo está hoy. Dicha razón tiene que ver con el denominado problema del Sol joven. A medida que el Sol envejece, se hace poco a poco más brillante debido a cambios en su constitución química.

Hace cuatro mil millones de años, habría sido un 30 por 100 más tenue de lo que es hoy, reduciendo drásticamente su efecto calentador sobre el lejano Marte. Esto estaría contrarrestado en parte por el calentamiento geotérmico, producido por la radiactividad y el calor almacenado procedente de la formación del planeta, y ambos efectos fueron mucho más fuertes en el pasado. Sin embargo, el flujo de calor geotérmico por sí sólo no compensaría el efecto del Sol joven, tenue, y hay que encontrar otras razones para un clima más tibio.
La manera más fácil de hacer un planeta más caliente es utilizando el efecto invernadero. Los gases invernadero tales como el dióxido de carbono actúan como un parasol, atrapando el calor del Sol cerca de la superficie del planeta. Hoy la atmósfera marciana es demasiado delgada para producir mucho calentamiento por efecto invernadero, pero ciertamente habría sido mucho más espesa durante los primeros mil millones de años. Como sucede con la Tierra, Marte adquirió una densa atmósfera inicial tanto por la degasificación del planeta como por el aporte de sustancias volátiles por parte de cometas, asteroides y planetesimales helados. Un CO2 abundante habría elevado la temperatura de modo espectacular.

Sin la presencia de agua, nunca se podrían formar estas figuras que vemos ahí. La erosión peroducida por el paso del agua, es inconfundible. En Marte había agua en abundancia que corría por su superficie, y, hoy, sabemos que sigue ahí congelada en la interperie y, no podríamos decir, en qué estado está en el subsuelo.
Aunque los científicos conjeturan que Marte debe de haber tenido mucho más CO2 en el pasado, no es fácil dar una cifra para ello. Primero hay que determinar dónde ha ido a parar el CO2. Con mucha probabilidad, la mayor parte de él se perdió en el espacio como resultado de impactos cósmicos masivos. La colisión de cometas grandes con planetas provoca erosión por impacto, que vacía la atmósfera. En el caso de Marte, el resultado final fue un aire poco denso, pero durante el propio período de bombardeo, la presión habría fluctuado de forma incontrolada. Los cálculos sugieren que Marte perdió el 99 por 100 a partir de entonces debido a procesos diversos. Si estos números son correctos, implican que Marte puede haber tenido en el pasado una presión atmosférica mil veces más alta que la actual, suficiente para elevar la temperatura por encima del punto de congelación y mantener incluso un extenso océano.

Existen marcas de las señales dejadas en las costas por los océanos de Marte. La subida del agua en bajamar y pleamar, dejaron señales inconfundibles de que, el océano estuvo allí. No hay dudas de que Marte tuvo alguna vez una atmósfera gruesa, puesto que las paredes de los cráteres producidos por impactos más antiguos han sufrido una fuerte erosión. Cráteres de menos de 15 kilómetro han sido completamente aniquilados. Por el contrario, los últimos cráteres apenas están erosionados. Tras datar el cambio, los investigadores creen que la atmósfera se redujo espectacularmente no mucho después del final del último bombardeo intenso, hace 3.800 millones de años.
La mayoría de las inundaciones catastróficas parecen haber ocurrido antes o aproximadamente en esa época, porque los canales de descarga están adornados por un montón de cráteres pequeños y bien conservados. Es la falta de erosión durante la mayor parte de la historia marciana la que ha mantenido sus cursos de agua extraordinariamente antiguos en una condición virginal. En la Tierra, ningún valle fluvial sobreviviría durante miles de millones de años.
Una vez que acabó el bombardeo, el dióxido de carbono de Marte siguió fugándose, por varias causas. Parte de él escapó al espacio, parte se disolvió en el agua o quedó absorbido en el regolito, y una gran cantidad puede haber quedado incorporada en carbonatos u otros minerales en las rocas. Sin algún proceso compensatorio, el CO2 hubiera sido engullido en muy poco tiempo. Probablemente, el calentamiento geotérmico invirtió algunos de estos procesos y devolvió parte del CO2 a la atmósfera.

Hasta el momento hemos podido enviar allí a distintos ingenios en forma de sondas espaciales y naves que, han hecho un buen trabajo y nos han acercado a la realidad de un planeta que, posiblemente, en su subsuelo, pueda contener algunas formas de vida. El Tiempo pasa, la técnica avanza y… ¿Que nos impedirá, más adelante, enviar robots más especializados para que, de una vez por todas investiguen si hay vida en el subsuelo del planeta?, o, todavía mejor, ¿Un viaje tripulado por astronautas una vez que logremos los conocimientos necesarios para asegurar sus vidas?
Durante unos cientos de millones de años puede haber habido una presión atmosférica moderadamente alta y un calentamiento asociado por efecto invernadero. Finalmente, sin embargo, el calor geotérmico desapareció, el reciclaje del CO2 flaqueó, y la presión atmosférica cayó en picado, produciendo el desierto liofilizado que vemos hoy en Marte.
El hecho de que parezca que algunos valles fluviales han sido excavados en fecha relativamente reciente sugiere episodios ocasionales de calentamiento. Una posible explicación procede de procesos de realimentación. Si un calentamiento geotérmico local o un estallido de vulcanismo llegasen a liberar repentinamente grandes cantidades de agua en la superficie, entonces un montón de dióxido de carbono disuelto se escaparía con ella. Esto, a su vez, elevaría la temperatura, con lo que se fundiría más agua y se liberaría más CO2. A medida que el agua fundida inundaba las bajas tierras congeladas, calentaría el regolito liberando aún más CO2. En total, podría haberse liberado en el planeta de esta forma incontrolada una cantidad suficiente de dióxido de carbono para crear temporalmente una atmósfera más densa con un calentamiento pronunciado por efecto invernadero.

¿Cuántas historias nos podrían contar los geólogos mirando está imagen?
Los Valles Marcianos que tardaron millones ede años en formarse, y, toda la orografía del planeta, nos habla de un pasado mejor, en el que el agua, estaba presente en abundancia y en los distintos estados en que son conocidos en la Tierra.
Otro mecanismo comodín concierne al movimiento del planeta. Marte tiene una órbita bastante excéntrica, y ninguna luna que pueda estabilizar su eje de giro. Habría habido veces en que condiciones favorables de los movimientos de rotación y orbital condujeran a un calentamiento solar considerablemente aumentado. En ocasiones, el eje de rotación podría haberse inclinado mucho, de modo que los polos recibieran más luz del Sol que las regiones ecuatoriales. Esto hubiera fundido los casquetes polares y producido un efecto invernadero en aumento. En el balance global, episodios repetidos de inundación, formación de océanos y glaciación, seguidos por largos períodos de inactividad, parecen más probables que el simple enfriamiento ininterrumpido.
Respecto a la posibilidad de vida, el hecho de que Marte estuviese caliente y húmedo hace entre 3.800 y 3.500 millones de años es altamente significativo, pues significa que Marte se parecería a la Tierra en una época en que la vida existía aquí. Esto ha llevado a algunos científicos a concluir que Marte habría sido también en esa época un lugar apropiado para la vida. Por sí misma, sin embargo, la presencia de agua líquida es sólo una parte de la historia. Lo que hace que las perspectivas de vida parezcan tan buenas es que Marte no sólo tiene agua líquida, sino también volcanes.
¿Hubo vida en Marte?

Lo que sí hay son heladas montañas y abundantes volcanes que nos hablan de su actividad del pasado.
La montaña marciana del Monte Olimpo se eleva 27 kilómetros sobre el macizo de Tharsis y tiene 550 kilómetros de diámetro. Medida por medida, es la montaña más grande de su tipo en el Sistema Solar, equivalente a amontonar siete montes Everets de la Tierra. La importancia del Monte Olimpo no está en su tamaño, sino en el hecho de que es un volcán. Donde se dan juntos volcanes y agua, pueden aparecer fuentes calientes: sistemas hidrotermales como los de la Tierra que posiblemente fueron un hogar para los primeros organismos. ¿Floreció también la vida microbiana en Marte hace 3.800 millones de años, quizá en alguna fuente burbujeante en la pendiente del Monte Olimpo, o en las profundidades de las rocas porosas por debajo de un mar marciano hace tiempo desaparecido?

La menor Gravedad de Marte, hace posible que sus montañas, sean inmensas, mucho mayores que las de la Tierra.
Hace cuatro mil millones de años, Marte aún resplandecía con el calor de su formación. La radiactividad calentaba la corteza. Los impactos cósmicos fundían la superficie. A medida que el planeta luchaba para deshacerse de este calor primordial, escupía lava de los volcanes a una escala masiva, creando inmensas llanuras de roca fundida similares a los mares de la Luna. A medida que la corteza se enfriaba lentamente, este vulcanismo declinaba continuamente: para la época que cesó el bombardeo intenso, estaba básicamente confinado a tres regiones principales: Tharsis, Elysium y Hellas. Si hay volcanes vivos hoy en Marte, no están manifestando ningún signo de actividad. Sin embargo, ha habido erupciones a lo largo de toda la historia marciana: por ejemplo alrededor del monte Olimpo dentro de los últimos mil quinientos millones de años, y cerca de Alba Patera en épocas tan recientes como hace quinientos millones de años. Puesto que es poco probable que Marte estuviera volcánicamente activo durante cuatro mil millones de años sólo para cesar su actividad en épocas relativamente recientes, parece razonable concluir que siguen existiendo algunos puntos calientes, probablemente en el subsuelo profundo.

Marte tiene regiones que nos son tan familiares como las de la Tierra
En el pasado remoto debe haber habido muchas oportunidades para que se formasen fuentes calientes alrededor de chimeneas termales, dada la abundancia de agua en el planeta. Hay clara evidencia de la interacción de agua y volcanes en los exámenes fotográficos. Muchas de las inundaciones fueron probablemente desencadenadas por lava que fundía el permafrost y el hielo del suelo, y se puede ver como algunos cursos de agua emergen claramente desde debajo de los flujos de lava. Los canales de desagüe se acumulan también alrededor de la región altamente volcánica de Tharsis. En otros lugares, densas redes de valles decoran los flancos de los volcanes.
Hay colinas de cima plana que se parecen a las tablas montañosas de Islandia, donde la lava ha rezumado desde debajo del hielo. Cordilleras de forma característica en Elysium llevan también la huella de una combinación de lava y hielo. Todo esto constituye una fuerte evidencia circunstancial de sistemas hidrotermales en el antiguo Marte, aunque todavía no han sido detectados depósitos minerales específicos, lo que sería un signo claro y evidente.

El Monte Olimpo
Mientras esperan nuevas misiones marcianas, los científicos de la NASA han estado ocupados en identificar puntos en la superficie del planeta donde podría haber tenido lugar actividad hidrotermal. La ladera del volcán Hadríaca Pladera parece un buen lugar. Aquí se encuentran muchos valles fluviales enmarañados que fluyen desde el borde de la antigua caldera, cruzados por un canal espectacular que emerge abruptamente a mitad de pendiente. Otro volcán, Apollinaris Patera, domina una región de aspecto singularmente brillante cerca del borde de la caldera, que podría ser un depósito mineral de fuente caliente. Un volcán similar en el área llena de cráteres conocida como Terra Cimmeria ha erosionado fuertemente las pendientes y está situado en el comienzo de un enorme curso de agua.
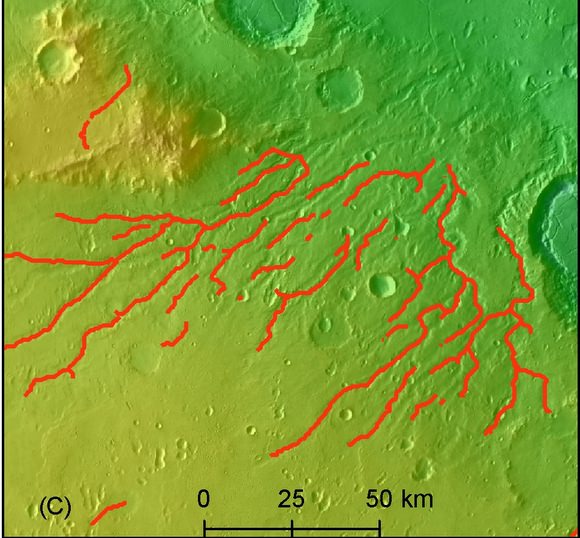
Muchos valles fluviales en Marte se dan en terreno caótico, donde hay grandes bloques de roca en masas revueltas. Se cree que esta topografía se formó cuando la roca fundida se introdujo en el hielo del suelo. Cuando el hielo se fundió, el agua fluyó haciendo que la tierra colapsara de una forma azarosa. Tales áreas serían un lugar perfecto para que aparecieran sistemas hidrotermales poco profundos.
Si, en efecto, la vida se asentó en una fuente caliente, quizá haya dejado restos fosilizados. Es probable que los fósiles marcianos hayan soportado las inclemencias del tiempo mejor que sus homólogos terrestres debido a la relativa falta de erosión climática. Futuras misiones de aterrizaje podrían buscar muestras para traer a la Tierra. Otros depósitos de fósiles potenciales incluyen valles fluviales, donde las inundaciones han podido arrastrar minúsculos organismos marcianos a las charcas estancadas, y la enorme grieta del Valle Marineris, donde estratos profundos han quedado expuestos. También tienen interés los lechos lacustres secos, en cuyos sedimentos se habrían podido depositar microbios. El cráter conocido como Gusev parece un candidato prometedor, puesto que un gran río desembocó una vez en él. Debe haber habido allí hace tiempo un lago profundo, con montones de sedimentos en el fondo.

La misión Pathfinder
El primer y pequeño paso siguiendo estos indicadores llegó en julio de 1977, cuando la misión Pathfinder depositó con éxito la primera nave espacial en Marte desde los tiempos de las Vikingo. Con su pequeño vehículo todo terreno Sojourner, la Pathfinder transmitió una gran riqueza de datos desde la boca de la llanura inundada Ares Vallis. En el terreno próximo a la nave espacial, hay esparcidas bolsa de rocas arrastradas por el torrente. Estos detritos podrían incluir fragmentos de un antiguo sistema hidrotermal, o incluso fósiles de microbios de la subsuperficie profunda llevados a la superficie con la inundación y transportados corriente abajo. Por desgracia, la Pathfinder no tenía capacidad de verificar estas conjeturas.
En septiembre de 1997, Mars Global Surveyor entró en órbita. Estaba diseñada para cartografiar la superficie del planeta con precisión en una escala de un metro y proporcionó una valiosa información sobre la historia hidrológica de Marte y los probables refugios para la vida. Hay Imágenes que nos hablan de una de la evidencia de una antigua orilla oceánica, charcas secas dentro de un cráter e incluso indicios de depósitos minerales asociados con sistemas hidrotermales, todo lo cual favorece las perspectivas de vida pasada.
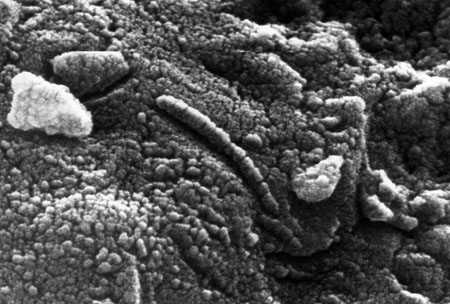
Hemos buscado la evidencia de la vida en aquel planeta y, no se descarta la idea de que, si seguimos insitiendo, la vida en Marte, aparecerá en forma fósil referida a la que estuvo presente en el pasado y, posiblemente, en los túneles y grutas que existen como vestigios de la rica vida volcánica del planeta, se encontrarán, formas de vida presente que vendrán a confirmar que, la vida, es algo natural en todo nuestro universo. Simplemente requiere del tiempo necesario y de las condiciones idóneas para su aparición.
emilio silvera